MAGNESIUM DEFICIENCY IN THE PATHOGENESIS OF DISEASE
Early Roots of Cardiovascular, Skeletal
and Renal Abnormalities
Goldwater Memorial Hospital
New York University Medical Center
New York, New York
1980
(include the word "jacket" to search only in this book)
| Jacket | Preface | Contents | Introduction (Chapter 1) |
Chapter: | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Appendix | Bibliography (A-D), (E-K),
(L-R), (S-Z) |
Part II: Chapter 5
MAGNESIUM DEFICIENCY IN THE PATHOGENESIS OF CARDIOVASCULAR DISEASES
5
Failure to Reduce Incidence of Ischemic Heart Disease by Lowering Blood
Lipids
Cardiovascular diseases continue to represent the major cause of morbidity and mortality in the developed countries, especially in young men, despite the considerable efforts and funds expended in the effort to explain and reverse the atherosclerotic process by studying and modifying fat intakes. Ischemic heart disease (IHD) is responsible for over 54% of all deaths in the United States (U.S. Dept. HEW, 1970); it is the major cause of death in most affluent communities (Editorial, Brit Med J, 1972). That the problem has increased during this century, particularly in young and middle-aged men, is indicated by two types of studies: (1) retrospective analyses of large numbers of necropsies (over 6000 each) in a large city hospital (Saphir et al., 1956) and from the Armed Forces Institute of Pathology (Pettyjohn and McMeekin, 1975); and (2) examination of the hearts of military men coming to autopsy in World War II (Yater et al., 1948, 1951; Moritz and Zamcheck, 1947), the Korean War (Enos et al., 1955), and the Vietnamese War (Macomber, 1971; McNamara et al., 1971; Wroblewski, 1971), and of victims of aircraft fatalities (Glantz and Stembridge, 1959). The study at Michael Reese Hospital in Chicago (Saphir et al., 1956) showed that there was an increase in frequency of coronary artery disease in subjects under 50 from 5.9% in 1920-1939, to 14.1% in 1940-1949, to 25.5% in 1950-1953. This did not take into account the almost twofold greater frequency of IHD in men than women. Pettyjohn and McMeekin (1975) found that 13% (816) of 6500 autopsied cases from aircraft accidents had been diagnosed as having had preexisting heart disease. Of those 816 cases (592 military and 135 civilian), 89.1% had coronary artery disease. Among the 380 men, 20-34 years of age, an upward trend was noted in the incidence of moderate to severe coronary artery disease from 1960-1964 to 1965-1969. (Too few autopsies were available for the 1970-1974 study for valid comparison.) In the studies of soldiers in the last three wars, startlingly high numbers of young men were found to have IHD. Yater et al. (1948) studied heart tissue from 866 American World War II soldiers, between 18 and 39 years of age, who developed IHD; 450 of these were examined at autopsy. From the incidence among soldiers, they estimated that the IHD death rate, per 100,000 men was less than 0.1 at 18-19, 1.0 at age 25-29, 3.4 at age 30-34, and 12.7 at age 35-39. Moritz and Zamcheck (1946) reported 115 sudden deaths from IHD in additional young soldiers. In the study of 300 American soldiers killed in Korea (Enos et al., 1955), 77% had histologic evidence of coronary disease; the average age was 22.1 years. The 1959 study of material from Air Force fatalities (Glantz and Stembridge, 1959) showed that 70% of 222 men of 20-44 had coronary disease, the highest incidence being in men 30-34, 35-39, and 40-44 years of age who had moderate to advanced arteriosclerosis in 32%, 26%, and 50%, respectively. There was a difference of opinion as to whether there was, indeed, a lower incidence of coronary artery disease among American soldiers killed in Vietnam (McNamara et al., 1971; Macomber, 1971; Wroblewski, 1971). Pettyjohn and McMeekin (1975) analyzed the factors contributing to the seeming decline in IHD incidence (McNamara et al., 1971) and attributed this finding to a difference in parameters used in classification of disease. The relative increase in mortality rates from IHD in the younger age groups has been confirmed by the international studies (International Workshop in Cardiovascular Disease, 1959-1969; Fejfar, 1974) even from groups employing measures to lower blood cholesterol levels (Fejfar, 1974).
As a result of the failure to prove that the incidence of deaths from IHD can be lowered by changing the fat intake of patients with the disease (Editorial, Brit Med J, 1972, 1976b; Stolley, 1972; Fredrickson, 1972; Fejfar, 1974), there has been revival of interest in the likelihood that adult cardiovascular disease has its roots in infancy, and that that is the time to change the fat in the diet (U.S. Dept. HEW, 1970; Glueck and Tsang, 1972; Glueck et al., 1972; 1974a). This approach is based on three findings: (1) the detection of fatty intimal streaks in arteries of infants and children (Duff and McMillan, 1951; R. Holman et al., 1958; R. Holman, 1961; Reisman, 1965; Strong and McGill, 1969); (2) the correlation of hyperlipidemia with increased risk of early arteriosclerosis (Gofman et al., 1950; Keys, 1956; Gertler et al., 1959; Berenson et al., 1974); and (3) the evidence that children of victims of early heart attacks often have hyperlipidemia (Tamir et al., 1972: Glueck et al., 1974b; H. Chase et al., 1974). Furthermore, large-scale screening programs have shown that three-year-old children already have cholesterol levels similar to those of young adults (Berenson et al., 1974). At present, it is considered feasible only to screen children with parenteral histories of early IHD (H. Chase et al., 1974; North, 1975; Laird, 1975). A general change of diet, so as to institute hypolipidemic regimens has been suggested (U.S. Dept. HEW, 1970). Altering the fat content of infants' diets substantially has been criticized because not all of the etiologic factors in arteriosclerosis are known, and because the results of the field trials have not yet proven that substituting unsaturated for saturated fatty acids will prevent coronary heart disease, even though they have lowered blood lipids (Stolley, 1972; Frederickson, 1972; C. Lowe, 1972; Levy et al., 1974; North, 1975; Laird, 1975). Furthermore, the potential risks of such diets remain to be ascertained (Foman, 1974; Schubert, 1973; Laird, 1975; Glueck et al., 1975/1977.
Thus, the need for searching out coronary-risk indicators persists (Blackburn, 1974). Changes in the musculoelastic layer of coronary arteries of infants and children are again being considered as the possible initial lesions in the atherosclerotic process (Neufeld, 1974; Danilevicus, 1974). The first visible changes in the internal elastic membrane, its splitting or fragmentation, are seen within a few days after birth (or in some cases in stillborn infants) and become more prominent in the first month of life (Bertelsen and Jensen, 1960; Bertelsen, 1961; Neufeld and Vlodaver, 1968, 1971, 1974; Neufeld, 1974). These are changes that have long been proposed as the first departure from normal, and that should be considered a manifestation of early arteriosclerosis and the basis for development of atherosclerotic lesions (Merkel, 1903; Jores, 1924; Minkowski, 1947; Fangman and Hellwig, 1947; Levene, 1956; Moon, 1957; Pizzagalli and Bertana, 1959; Bertelsen, 1961; Kaunitz, 1961; Gillot, 1962).
Many factors contribute to the metabolic abnormalities that lead to different blood, arterial, and cardiac biochemical, functional, and histological changes that represent aspects of the complex of cardiovascular diseases. Vitamins B6 and E have been suggested as protective against arteriosclerosis. Vitamin B6 has been suggested by Rinehart and Greenberg (1949, 1951, 1956), Moon and Rinehart (1952), Moon (1957, 1959), Boxer et al. (1957), Hass (1961), and Levene and Murray (1977). Vitamin E has been suggested in peripheral disease, e.g., intermittent claudication, by Livingstone and Jones (1958), Haeger (1968, 1973), Larsson and Haeger (1968), and Williams et al. (1971); in thrombotic disease by Zierler et al. (1948), Ochsner (1951), Suffel (1956), and Kawahara (1959); and in the controversial use in heart disease by Vogelsang et al. (1947; publications of the Shute Institute). Pyridoxine/blood- and tissue-lipid interrelationships have long been known (Birch, 1938; Medes and Keller, 1948; Schroeder, 1955; Shah et al., 1960; G. Emerson et al., 1960; Lupien, 1968), and combinations of the vitamins, sometimes with A (Hammerl and Pichler, 1960) proposed. Vitamin C has been shown to lower plasma cholesterol levels, and by inference atherosclerosis (Spittle, 1970, 1971; Anderson et al., 1972). However, hypercholesterolemia has been produced in rats by supplements of vitamin C equivalent to excesses of less than one gram over that in the diet, an effect attributed to ascorbic acid induced production of high zinc/copper ratios (Klevay, 1977). Thiamine deficiency has been shown to increase the hepatic synthesis of lipids by rats; hypertriglyceridemia of magnesium deficiency develops in the presence of adequate or excess thiamine, but not in double deficiency (Itokawa et at., 1973). Excess vitamin D increases arteriosclerosis both in experimental animals and in man-infants, children, and adults. In his epidemiologic correlation of only slightly higher than recommended intakes of vitamin D with increased incidence of myocardial infarction, Linden (1974b) suggested that the hypocholesterolemic effect of vitamin A might protect against the hypercholesterolemic action of vitamin D. Additional studies confirm that experimental A deficiency increases both atherosclerosis and cholesterol blood levels (Bayer et al., 1972; Bonner et al., 1973). In 1962, I. Clark and Bassett showed that vitamin A decreased other manifestations of vitamin D toxicity: osteolysis and renal and arterial calcinosis. As early as 1939, Reed et al. reviewed the data on vitamin D toxicity and reported that in the absence of vitamin A, the lesions of hypervitaminosis were worse.
Several of these vitamins are of interest in this presentation because they affect magnesium metabolism, influence the response to magnesium, or affect magnesium requirements. For example, vitamin B6 deficiency causes arterial lesions (Rinehart and Greenberg, 1949, 1951) very much like those of magnesium deficiency (Hass, 1961), and gestational B6 deficiency has been blamed for the elastica damage of neonatal arteriosclerosis (Levene and Murray, 1977), in which magnesium deficiency is implicated in this book. Interrelationships of magnesium and pyridoxine metabolism have been reviewed by Durlach (1969b). The response of B1-deficient animals (Zieve et al., 1968a,b; Zieve, 1969) and man (Zieve, 1975) is magnesium dependent. The cardiovascular lesions of vitamin D excess might be partially implemented by magnesium loss. Vitamin D increases magnesium loss by increasing its renal excretion relative to its absorption (Hanna, l961b; Richardson and Welt, 1965; Wallach et al., 1966). Infants and children with hypervitaminosis D, who develop hypercalcemia, the supravalvular aortic stenosis syndrome (SASS), and other stenotic and hypoplastic lesions of the greater arteries, also develop peripheral arterial lesions (including atheromata and calcinosis), hypercholesterolemia, and hypertension. Since vitamin D excess causes calcium retention as well as magnesium loss and high calcium/magnesium plasma ratios have produced increased arterial resistance (Review: Haddy and Seelig, 1976/1980, and infra vide), the combination of low magnesium stores at birth and high vitamin D and calcium intakes in infancy can be responsible for several metabolic and histologic aberrations leading to cardiovascular diseases.
The similarity of the arterial lesions, and of the microfocal myocardial necrosis seen in the infants, to those produced experimentally by essentially "pure" magnesium deficiency (Seelig and Haddy, 1976/1980) has suggested magnesium deficiency during gestation and infancy. Furthermore, magnesium deficiency, even in the absence of hypervitaminosis D, has been shown to cause abnormal changes in blood lipids (Review: Seelig and Vitale, 1971/1973).
Epidemiologic data point both toward magnesium as a protective factor against sudden death from ischemic heart disease and toward even slight to moderate excesses of vitamin D as a risk factor in hyperlipidemia and myocardial infarction (Review: Linden, 1975/1977). Experimental data demonstrate that magnesium is protective against several models of myocardial disease, including those caused by hormonal and nutritional imbalances, stress, and hypoxia (Reviews: Seelig, 1972; Seelig and Heggtveit, 1974).
It is important to note that the proposal that magnesium deficiency is a contributory factor in cardiovascular disease does not negate the role of high fat intakes (which interfere with magnesium absorption). Also, the theories implicating the fatty streak as an early infantile atherosclerotic lesion do not preclude the theories that elastica degeneration is one of the earliest arterial lesions. Lipid droplets are seen in conjunction with damaged elastica (Duff and McMillan, 1951) and have been correlated with elastica degeneration (Pickering, 1963; Zugibe, 1963). Furthermore, elastica degeneration predisposes to lipid deposition (Kramsch et al., 1970, 1971). The papers that stress the changes in the musculoelastic layers of the arteries of infants as the earliest signs of arteriosclerosis support the premise that magnesium deficiency during the perinatal period, and factors that increase magnesium loss then and in early childhood, can contribute to the pediatric origins of cardiovascular disease, since comparable changes are seen in magnesium deficiency (Review: Seelig and Haddy, 1976/1980.
5.1. Magnesium and Lipid Interrelationships
5.1.1. Influence of Fat on Magnesium Retention (Man)
Because excess (saturated) fat has been considered a major contributory factor in atherosclerosis and ischemic heart disease, and experimental and epidemiologic studies implicate magnesium deficiency (infra vide), evidence of interrelationships between fat and magnesium is considered first.
5.1.1.1. Dietary Fat and Magnesium Balance
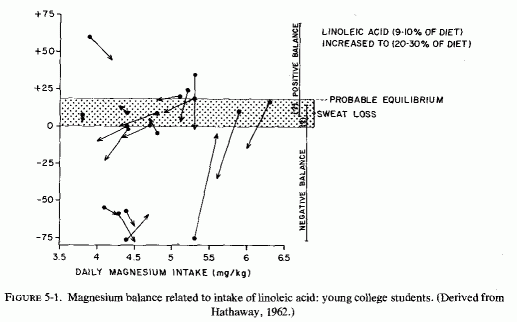
Evidence was obtained early that diets rich in fat interfere with magnesium absorption. In 1918, Sawyer et al., performed metabolic balance studies with 2 boys, 5 and 8 years of age, in which they explored the effects of fat intake on retention of calcium and magnesium. Even though their magnesium and calcium intakes were lower than in their normal diet, increasing the fat intake resulted in their excreting more of the divalent cations in both feces and urine. In a study of mineral balance of 4 young women on controlled magnesium-rich diets (800 mg/day), substitution of butter for vegetable fat resulted in retention of more magnesium (Bogert and Trail, 1922). On lower intakes of magnesium (320-350 mg/day), 6 young men given controlled diets containing 10-30% linoleic acid tended to be in negative magnesium balance while on the fatty-acid-supplemented diet (Irwin and Wiese, 1961). More extensive metabolic balance studies confirmed the interference by unsaturated fatty acids with magnesium retention in 19 young men on typical American intakes of magnesium, averaging 300-400 mg/day or 3.8-6.3 mg/kg/day (Hathaway, 1962). Magnesium, calcium, and phosphorus balances were calculated for ten 5-day periods, during which the diets were supplemented with linoleic acid at 9-10% and a subsequent increase in fatty acid to 20-30%. Most of the subjects were in probable magnesium equilibrium on the low-fat diets, considering balance to fall between 0 and + 18 [the sweat loss of magnesium, that averages 18 mg/day (Seelig, 1964) not being allowed for in the figures given (Hathaway, 1962)]. When the fat intake was increased, although the magnesium intakes remained approximately the same, most retained less magnesium (Fig. 5-1). Only 1 of the 19 men remained in strongly positive magnesium balance on the high fat intake. Of 11 who were in ± magnesium balance during the low fat intake, 6 showed essentially no change on high fat, and 5 went into negative magnesium balance. Two, who were in slightly positive magnesium balance on low fat, dropped to no retention on high fat. Of five who were in strong negative balance on the low-fat diet, four continued to lose substantial amounts of magnesium when the fat intake was increased, and one lost less. Thus, 11 of the 19 subjects lost more magnesium on the high- than on the low-fat diets.
In a 5-day metabolic study of five normal medical students given a liquid diet, such as that given to preoperative peptic ulcer patients (that delivered 138 g of fat, predominantly from whole milk and cream, and that provided only about 200 mg of magnesium daily), there was an average daily loss of 16 mg of magnesium (Macbeth and Mabbott, 1964). This amount of dietary fat is equivalent to that of the typical American diet, comprising about 40% of the daily calories (de los Rios, 1961). This liquid diet differed from that given to the young men on the linoleic acid diet (Hathaway, 1962) in that the ratio of calcium to magnesium was 9:1, rather than 2:1.
Metabolic balance studies in rats on low- and high-fat diets have also shown that increasing the fat intake decreases the amount of magnesium absorbed from the gut (Olson and Parker, 1964; Tadayyon and Lutwak, 1969).
5.1.1.2. Steatorrhea and Magnesium Loss
Diseases that interfere with fat absorption, thereby resulting in high concentrations of fat in the intestinal lumen, interfere with magnesium absorption in several ways. Insoluble complexes of the fats and magnesium and calcium prevent their transit across the intestinal membranes, in conditions such as celiac disease or sprue, and in the steatorrhea that develops after gastrointestinal resections. [Bowel resections or bypasses further contribute to magnesium depletion by shortened transit time, decreased absorption area, and decreased enzymatic lipolysis (Opie et al., 1964).]
5.1.1.3. Dietary Fat and Blood Lipids (Man)
Despite the evidence that fat interferes with magnesium absorption, and that steatorrhea has caused hypomagnesemia and acute and subacute magnesium-depletion syndrome, the short-term studies of the effect of high fat intakes on serum magnesium have shown no effect For example, Macbeth and Mabbott (1964) who found that young men on a "Sippy-like" ulcer liquid diet for five days, who were in negative balance, maintained normal serum magnesium levels (1.9 ± 0.34 mEq/liter) Studies with older (42-62 years of age) schizophrenic patients on diets delivering 34, 65, and 134 g of fat (each patient given each of the diets for two-week periods in different sequences) showed no change in serum magnesium levels on the different diets though serum cholesterol levels fell on the low-fat diets (de Los Rios, 1961).
The epidemiologic studies of residents of hard- and soft-water cities, which consider the fat intakes, the magnesium and calcium content of the water supplies, and serum lipid and magnesium and calcium levels (Bierenbaum et al., 1973), provide interesting insight into the protective effect of hard water against sudden death from ischemic heart disease (supra vide). Comparison of these parameters in hard-water American and English cities (Omaha, Nebraska and London) with soft-water American and Scottish cities (Winston-Salem, North Carolina, and Glasgow) provides data that implicate the cations more than the fat ingested in the substantially lower cardiac death rates in Nebraska and in London than in the soft-water cities of southeastern United States and Glasgow (Review: Seelig and Heggtveit, 1974, and supra vide). For example, there was no significant difference in the percentages of those in Omaha and Winston-Salem (51.5% and 47.4%) who ingested diets high in fat, and in their serum cholesterol, triglycerides, and phospholipids. The tested residents of Glasgow, 72.8% of whom ate diets high in fat, had essentially the same serum lipid levels as did the tested London residents, only 28.2% of whom ate high fat diets. Nonetheless, the serum cholesterol and triglyceride levels of Glasgow residents were the lowest of all the four cities. Also, residents of both hard-water cities had higher serum cholesterol than did those of theft paired soft-water cities (p < 0.05). In evaluating the comparable serum levels of magnesium in both American cities, but the significantly higher serum magnesium in London than Glasgow, the noted common use of water softeners in Omaha (but not in London) should be considered. Residents of both American cities had significantly higher serum calcium levels (10.37 in Omaha, 9.59 in Winston-Salem) than did those in Britain (8.57 in London, 8.7 in Glasgow). The possibility that this is a reflection of more milk and vitamin D ingested by adults in the United States than in Britain should be considered, since the calcium content of London water was 2-3 times as high as that in the American cities.
With these data in mind, it is not surprising that there has been disagreement as to correlation of serum magnesium and cholesterol levels in patients with cardiovascular disease, or in populations at different risk.
5.1.1.4. Serum Magnesium and Cholesterol Levels in Cardiovascular Patients and High-Risk Populations
Bersohn and Oelofse (1957) and Bersohn (1958) correlated the lower serum cholesterol and slightly higher magnesium levels in Bantus than in white South Africans, with the lower incidence of arteriosclerosis and the higher dietary intake of magnesium of the Bantus. They analyzed the serum magnesium levels of Europeans with low to high serum cholesterol levels and found that, although there was overlap of serum magnesium values, the mean magnesium level of those with low serum cholesterol (mean = 170 mg/100 ml) was higher (1.7 mEq/liter) than was that of patients with hypercholesterolemia (cholesterol: 310-586 mg/100 ml; magnesium: 1.4 mEq/liter). In an Australian study comparing serum cholesterol and magnesium levels in several groups of aborigines and Europeans, Charnock et al. (1959) confirmed the lower serum cholesterol levels of the aborigines (who have a low incidence of cardiovascular disease) than of the Europeans and found significant differences (p = 0.001) between serum magnesium levels of the aborigines (1.7 mEq/ liter) and the Australians living in Adelaide (1.2 mEq/liter). Another group of Australians, living in a northern area (Alice Springs) one thousand miles away, had high mean serum magnesium levels (1.9 mEq/liter) and the highest mean serum cholesterol (314 mg/100 ml) of all the groups tested. Thus, the correlation between magnesium and cholesterol levels was not consistent. (The nature of the water of Alice Springs was not given.) It was interesting that there was no difference in serum magnesium (1.3 mEq/liter) and cholesterol levels (286; 281 mg/100 nil) in ischemic heart disease patients and age-matched European controls in Adelaide.
D. F. Brown et al. (1958), noting the report by Bersohn and Oelofse (1957) (supra vide) and that by Malkiel-Shapiro et al. (1956) that parenteral administration produced clinical improvement and lowered β-lipoprotein levels in patients with myocardial infarction (MI), studied serum magnesium-lipid relations in MI patients and in middle-aged controls. They found no correlation between serum magnesium and lipid levels, and no significant difference between the patients and the controls. Similar negative findings have been reported by others in studies of patients with cardiovascular disease and hyperlipidemia (Hyatt et al., 1966; Murnaghan et al., 1969; Rotman et al., 1971/1973).
On the other hand, Jankelson et al. (1959), who compared serum magnesium and lipid fractions of atherosclerotic patients and controls, found that although total cholesterol levels were the same in both groups, there were differences in magnesium and lipoprotein levels. The avenge serum magnesium level was 1.4 mEq/liter in 23 atherosclerotic patients and 1.6 in 12 healthy controls (in third and fourth decades of life). The β-lipoproteins were 11.5 in patients and 8.5 in controls; the α-lipoproteins were 5.3 in patients and 2.2 in controls. Six of the atherosclerotic patients were alcoholics; all had normal cholesterol levels, but 4 had high β-lipoprotein and all had higher than control α-lipoprotein values; 4 had very high levels. There was not good correlation, however, of low serum magnesium levels with high lipoproteins. Three with arteriosclerotic heart disease, and/or cerebral thrombosis, respectively, had hypomagnesemia (0.7, 0.7, and 1.3 mEq/liter) and hyper-β-lipoproteinemia (8, 13.4, and 9). But 2 with comparable disease and high β-lipoproteins (12.5 and 14.3) had normal serum magnesium (1.7 and 2.0 mEq/liter). One with cerebral thrombosis had normal magnesium and lipid levels. High serum cholesterol levels (288 mg/100 ml) and low serum magnesium levels (1.5 mEq/liter) were seen in 25 patients with acute MI a week after the infarction, as compared with the avenge levels in 50 controls (cholesterol: 210; magnesium: 2.1 mEq/liter) and in 15 old MI cases (cholesterol: 238; magnesium: 1.9) (Nath et al., 1971/1973). The magnesium levels rose during the next two weeks to normal (1.9 mEq/liter). Patients with angina pectoris, in this series, had high cholesterol levels (278) but normal serum magnesium values (2.0 mEq/liter). Rangam and Gupta (1961) found that among 44 patients with hypercholesterolemia, 80% had hypomagnesemia; among 52 with high lipid phosphorus levels, 75% had low serum magnesium levels. Those with normal cholesterol levels, however, also had a high incidence (54%) of hypomagnesemia in this series.
A brief abstract reports highly significant (p = .001) correlations between magnesium and high cholesterol and low-density lipoproteins in a survey of 32 random subjects 40-60 years of age (Mondschein, 1974). Over half of the magnesium values were below the normal range; none was above.
5.1.1.5. Clinical Use of Magnesium in Cardiovascular Disease with Hyperlipidemia
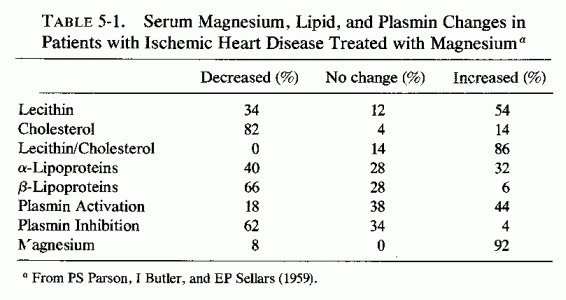
Many of the attempts to determine whether high serum cholesterol levels correlated with low magnesium levels in patients with cardiovascular disease derived from the clinical reports that parenteral magnesium administration was of value in treating patients with myocardial infarcts (MI), coronary insufficiency, and/or peripheral arteriosclerosis. Malkiel-Shapiro et al. (1956) first reported lowering β-lipoproteins in patients with coronary insufficiency, with the use of intramuscular (i.m.) MgSO4 begun at the time of an acute attack of coronary thrombosis or during acute coronary insufficiency. The regimen employed by Malkiel-Shapiro (1958) for 25 years involved deep i.m. injections of 0.5-2.0 ml of 50% MgSO4 on alternate days, and at longer intervals (to twice weekly) as the condition improved. Patients who had recently recovered from an MI or who suffered angina of effort were usually given 12 i.m. injections of MgSO4 at 5-day intervals, which was repeated after 4-6 months if they had benefited. These physicians stated that patients with more advanced disease seemed to have the most striking improvement. In the earlier studies (Malkiel-Shapiro et al., 1956), patients on magnesium therapy were given no anticoagulants. Subsequently, concurrent use of small doses of heparin (5000 units daily) and i.m. MgSO4 (0.5 ml 50% solution) once weekly were found useful in the maintenance of patients who have recovered from an acute MI (Malkiel- Shapiro, 1958; Malkiel-Shapiro et al., 1960). After the 1956 report of this group, clinical trials were undertaken in South Africa (Teeger, 1958; Agranat 1958; Marais, 1958) and Australia (R. Parsons, 1958; R. Parsons et al., 1959, 1960, 1961), and there was clinical verification of much that had been claimed. Agranat (1958) reported a 44% improvement rate with the use of MgSO4 injections to patients with chronic IHD. R. Parsons (1958) reported briefly that 3 injections of 1 ml 50% MgSO4 weekly for a month to IHD patients resulted in reversal of low lecithin/cholesterol ratio, lowering of the β-lipoprotein levels with elimination of the pre-β- band, and reduction of plasmin inhibition. In a detailed report, R. Parsons et al. (1959) described treatment of patients with angina but no ECG evidence of MI, and of patients with ECG evidence of MI with or without angina. They found that 2-ml doses of 50% MgSO4 given i.m. every 5 days (until 12 doses were given), were more effective than were the 1-ml doses. Patients with acute MI were also given heparin for the first 3 days of treatment. Comparison of the results of this regimen with that obtained the previous year when only anticoagulants were used were striking. Of over 100 patients given the magnesium therapy, one-third of whom had had acute MI, there was only one death. Among almost 200 patients treated with anticoagulants alone, 60 died. The biochemical changes (Table 5-1) show the improvement in lecithin/cholesterol ratio, the decrease-particularly in β-lipoproteins-and in plasmin inhibition produced by the magnesium therapy. In 1960, Parsons et al. published confirmation of the observation (Malkiel-Shapiro, 1958) that combination of low dosage heparin with i.m. magnesium therapy was even more effective in speedily reducing β-lipoproteins and total lipids to normal levels. They recommended that patients with acute MI should be given heparin (15,000 units every 6 hours for 3 days), with an initial dose of 2 ml 50% MgSO4 i.m. Then low-dosage (5000 units) heparin and 2 ml 50% MgSO4 were given three times weekly for 6 weeks, and once weekly subsequently.
Application of this regimen to patients with angina, MI, or peripheral arterial disease (incipient gangrene, ischemic leg ulceration, Raynaud's disease, and intermittent claudication) has been reported to produce clinical improvement and to lower serum cholesterol levels (S. Browne, 1961, 1963, 1964a,b). Savenkov et al. (1971) have also reported that treatment with a preparation containing magnesium adipate and magnesium nicotinate (in tablet or ampoule form for i.v. or i.m. administration) has been useful in the treatment of 54 patients with coronary, cerebral, and (in 21 cases) peripheral atherosclerosis. Treatment was given for 20 days parenterally (18 patients), orally (20 patients), or parenterally for half the course, followed by oral administration (16 patients). The clinical response was considered good in 22 instances, satisfactory in 16, and effective in 10. The total serum cholesterol was obtained in 41 patients (average = 284 ± 16.9 mg/100 ml). The level decreased in 29 patients, did not change in 9, and rose by 24 mg/100 ml in 3. In the entire group, there was an average decrease of 17.5%.
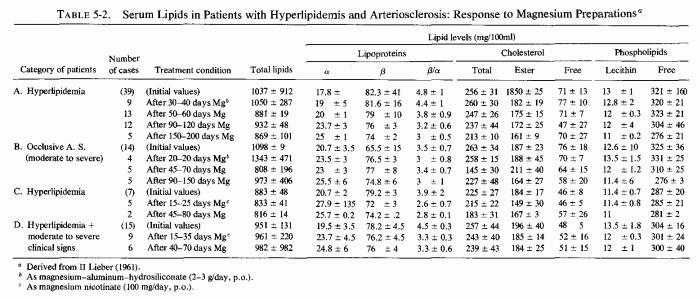
A magnesium-aluminum-siliconate preparation was given in fairly high dosage (2-3 g/day) to hyperlipemic patients without (Table 5-2A) and with (Table 5-2B) moderate to severe manifestations of cerebral, coronary, or peripheral arterial disease (Lieber, 1961). The most reduction was in the esterified cholesterol fraction, an interesting finding in view of the high levels of cholesterol esters in magnesium-deficient dogs (Kruse et al., 1933) and rats (Savoie and Delorme, 1976/1980). The ratio of β/α-lipoproteins fell increasingly with the duration of administration of the magnesium preparation, and to a greater degree in group A than group B. The low-dosage magnesium nicotinate preparation was given to patients with lesser degrees of hyperlipidemia, and produced less striking changes, but in the same direction (Table 5-1C, D) (Lieber, 1961). The tendency toward magnesium-induced decreased β-lipoprotein levels is reminiscent of comparable findings in magnesium-supplemented rats on atherogenic diets (infra vide).
A brief abstract of a long-term (19-month) double-blind study of 35 patients given either oral MgCl2 and KCl (1 mEq/kg/day) or placebo, reported that serum β-/α-lipoproteins were 10% lower in the treated group than in the placebo group (Haywood and Selvester, 1962). The dose-limiting effect of side effects was considered a likely explanation of the failure to reduce the lipids further. Smaller doses of magnesium given in complexed or chelated form were reported to lower the elevated β-fraction somewhat, but not to lower the total cholesterol levels; when the magnesium was stopped the β-fraction rose to pretreatment levels (A. Steiner, 1962, 1963). In the latter study, β-vitamins were also given.
Rademeyer and Booyens (1965), having shown that the maize meal dietary constituent of the Bantus had a hypocholesterolemic effect in rats, which they attributed to its high magnesium content and to its interference with fat absorption, demonstrated that supplementation with maize meal of diets of hyperlipemic whites raised their serum magnesium and lowered their serum cholesterol levels (Booyens et al., 1966).
The conflicting clinical reports as to the relationships of serum magnesium and lipid levels, and their meaning in cardiovascular disease, call for evaluation of magnesium-lipid interrelationships in experimental atherogenesis. In particular, because of the poor correlation between serum lipid levels and protection against heart disease in hard-water areas (Bierenbaum et al., 1973), and because magnesium administration has been claimed to be beneficial in overt clinical cardiovascular disease, animal data deserve careful scrutiny.
Among the numerous studies of atherosclerosis induced by fat, cholesterol, and cholic-acid-loading of animal diets, few have included determination of magnesium levels. The early magnesium-deficiency studies by Kruse et al. (1933) showed that young dogs on an atherogenic diet low in magnesium (.08% of diet) and containing butter fat (8% of diet), exhibited no change in total blood lipids, little or no change in free cholesterol, a drop in fatty acids, but a substantial rise in esterified cholesterol. As the magnesium level dropped, the percentage of esterified cholesterol rose. The young dogs on a magnesium-free, but otherwise well-balanced, diet, which delivered no lipid other than corn oil, were found by Vitale et al. (1961) to develop neither elevation of blood cholesterol nor atheromatous plaques. Bunce et al. (1962a,b) demonstrated that increasing the magnesium intake from 80 to 180 ppm, in dogs fed 20%-animal-fat diets, prevented the aortic lesions seen in dogs on the lower magnesium intake, but allowed for a slight further rise in serum cholesterol.
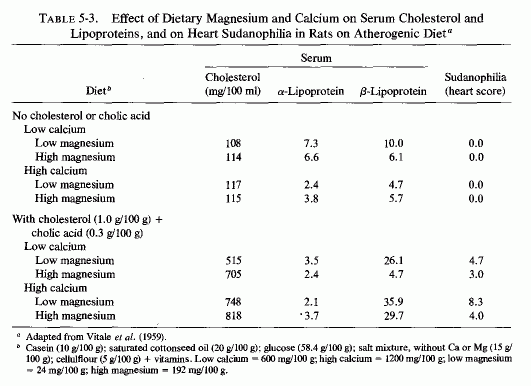
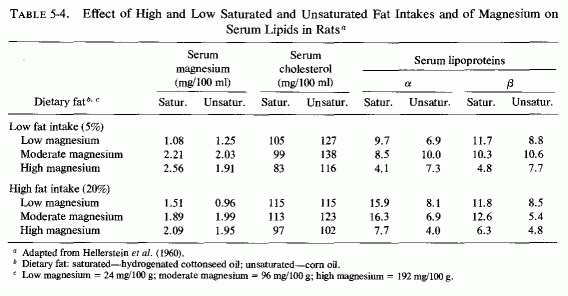
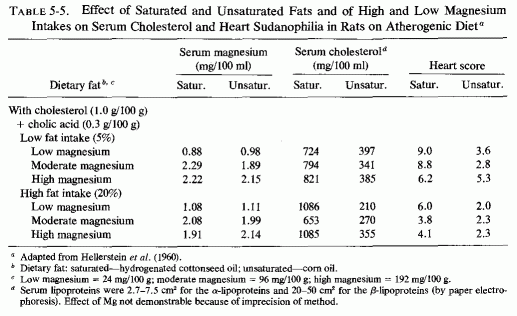
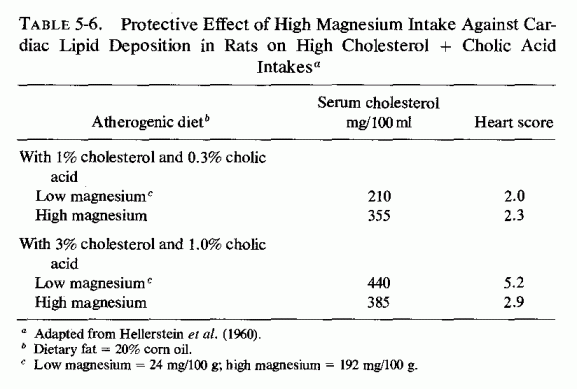
The atherogenic diet fed to rats (Vitale et al., 1957a,c,d,e; Hellerstein et al., 1957, 1960) produced marked hypercholesterolemia (639-808 mg/100 ml) that was not lowered by increasing the magnesium intake, even though early arteriosclerotic lesions were diminished (Vitale et al., 1957d,e; Nakamura et al., 1960). In rats on the atherogenic diet, also high or low in protein (Vitale et al., 1957c) or calcium (Vitale et al., 1959), increasing the magnesium content caused a further rise in serum cholesterol. A high intake of both magnesium and calcium, reduced the sudanophilia of the hearts to 4.0 from the high value of 8.3, but exerted little influence on serum lipids (Table 5-3). Increasing the magnesium intake of rats on low calcium intake substantially lowered the β-lipoproteins. A high magnesium intake slightly lowered the serum cholesterol and more profoundly lowered the lipoproteins of rats on high and low fat intakes, whether the fats were saturated or unsaturated (Hellerstein et al., 1960: Table 5-4). No cardiac sudanophilia developed, unless cholesterol and cholic acid were added to the diet. The markedly elevated plasma cholesterol, seen in rats also given cholesterol and cholic acid, was actually increased on the higher Mg intakes, although the cardiac lipid deposition in the rats on saturated fats and high Mg was reduced. Altering the Mg intake did not notably affect the lesser heart scores of rats on high intakes of unsaturated fat (Table 5-5). The elevation of heart scores of rats on low unsaturated fat diets when their magnesium intake was increased requires elucidation. Vitale et al. (1959) and Hellerstein et al. (1957, 1960) suggested that magnesium might protect against lipid deposition in the cardiovascular system by means of its effect on lipoprotein metabolism. They demonstrated that further increasing the intakes of cholesterol to 3% and cholic acid to 1%, of rats on 20% unsaturated fat, increased serum cholesterol levels only slightly (to 440 mg/100 ml), but increased the heart scores of rats on low magnesium intake to 5.2. High magnesium intake protected against this increased heart score (Table 5-6). Increasing the magnesium intake of rats on atherogenic diets, given alcohol or water to drink, also resulted in higher serum cholesterol levels, but less cardiovascular sudanophilia (Vitale et al., 1957a). Nakamura et al., (1960, 1966) showed that the long-term feeding of 192 mg/100 g of magnesium to rats on this atherogenic diet produced an early increase in serum lipids that fell only gradually within the year-long observation, but a significant decrease in arterial lipid deposition was evident within two months on the magnesium-supplemented diet.
In contrast to the results in the foregoing studies with hypercholesterolemic semisynthetic diets, the high blood cholesterol produced in rats fed whole milk (containing 4 g butter fat/100 ml milk) alone or with added cholesterol, was corrected by adding MgSO4 to the diet (Mullick and Kakkar, 1963). It seemed possible that formation of insoluble compounds of the milk fat and magnesium might have prevented absorption of the excess fat. However, in another report, magnesium salt given intramuscularly also lowered the serum cholesterol (Kakkar and Mullick, 1963).
Rademeyer and Booyens (1965) explored the effect of butter fat versus sunflower-seed oil on the serum magnesium and cholesterol levels of rats fed a semisynthetic low magnesium diet similar to that used by Vitale's group (supra vide). They found that the addition of 25% butter fat to the diet lowered the serum magnesium from 3.3 to 2 mEq/liter and raised the serum cholesterol from 65.8 to 81.6 mg/100 ml over a 4-week period (p = 0.001). The serum magnesium did not fall on addition of 25% sunflower-seed oil, nor did the serum cholesterol rise. An equal amount of meat-fat drippings caused a lesser fall in serum magnesium than did the butter fat, and lesser but significant rise in serum cholesterol. Substituting sunflower-seed oil for butter in the group that had been fed the butter-supplemented diet for 4 weeks affected neither the depressed serum magnesium nor the elevated serum cholesterol, but substitution of maize meal for glucose caused a rise in magnesium and a fall in cholesterol within a week. Maize meal (a major dietary constituent of Bantus) was used in this study in an effort to determine why Bantus have a lower serum cholesterol and higher serum magnesium level, as well as a lower incidence of arteriosclerosis than do South African whites (Bersohn and Oelofse, 1957).
Hungerford and Bernick (1976/1980) have recently reaffirmed the lack of alteration of plasma magnesium in rats on synthetic atherogenic diets, and elucidated the histologic arterial changes produced by an atherogenic or magnesium-deficient, or combined high-fat low-magnesium diet. They showed the further increase in serum cholesterol produced when rats on atherogenic diets were also magnesium deficient.
Rabbits on a hypercholesterolemic diet for 24 weeks showed a sharp drop in serum magnesium (and calcium) at 6 weeks. The hypomagnesemia persisted for 6 more weeks and then tended to rise (Rangam and Gupta, 1961). Intravenous MgSO4 injection (2 ml 5% solution) to such rabbits was found to lower serum cholesterol for 48 hours (Rangam and Gupta, 1962). Magnesium deficiency intensified the deposition of fat in the aortas of rabbits on atherogenic diets, lowered the level of serum triglycerides significantly (p = 0.05), but exerted little effect on total serum cholesterol (Nakamura et al., 1965). Magnesium supplementation had little effect on serum aorta lipid levels in rabbits in one study (C. Adams et al., 1964). Neal and Neal (1962) found higher serum phospholipid and triglyceride levels in rabbits on atherogenic diet when their drinking water contained magnesium than when they were given distilled water to drink, but they had less atherosclerosis when they were magnesium supplemented. Another group confirmed these observations. They found that administration of magnesium (as Mg Na2 EDTA) had little effect on the hyperlipidosis of rabbits on atherogenic diet but reduced formation of atheromatous plaques (McCann et al., 1962; Wartman et al., 1967). The magnesium-deficient cebus monkeys on atherogenic diets, reported by Vitale et al., (1963), showed both elevated serum cholesterol values and marked intimal lipid deposition in the aorta, not seen in controls. A study of the response to [3H]cholesterol given intravenously to magnesium-deficient and control rats, showed that the tagged cholesterol was taken up and subsequently released more rapidly by the liver of magnesium deficient than control rats. As a result, there was an initially greater drop in serum [3H]cholesterol and a greater subsequent rise in the magnesium-deficient rats; they also exhibited extracellular [3H]cholesterol between the elastic lamellae and the smooth muscles in the aorta (Schmalbeck et al., 1972).
The Mg- and KCl-free diet, containing animal fat, vitamin D, and sodium phosphates, which was contrived by Sos et al. (1964a,b,c) to be cardiopathogenic in several species, produced elevated serum cholesterol levels (Review: Seelig and Haddy, 1976/1980). A similar diet, designed to be thrombogenic, but that also produced cardiac necrosis in rats (Savoie, 1972a,b, 1975; Savoie et al., 1973), produced a substantial rise in blood cholesterol levels, particularly in the esterified form (Savoie and Delorme, 1976/1980), a finding that recalls the early observation by Kruse et al. (1933) in dogs. Blood phospholipids were also increased, and blood magnesium levels were lowered. Magnesium supplementation of the atherogenic or of the thrombogenic diet exerted little effect on most of the blood lipid fractions, raising some further and lowering some slightly but none to normal levels (Savoie and Delorme, 1976/1980). It is noteworthy that hypocholesterolemic agents (clofibrate, nicotinic acid, and conjugated estrogens) exerted no protective effect against the nonocclusive suppurative cardiac necrosis produced when Na2 HPO4 was added to the hyperlipemic thrombogenic diet. Only MgCl2 was completely protective (Savoie, 1972b). Further work showed that the sodium phosphate addition accentuates the hypokalemia of the thrombogenic diet, but produces hypomagnesemia, and lowers the cardiac magnesium levels. More recently, Savoie and Delorme (1976/1980) found that the thrombogenic diet increased lipoprotein lipase activity, an effect not influenced by magnesium. On the other hand, the added phosphate lowered the cardiac lipase activity, and magnesium raised it, with resultant elevation of cardiac free fatty acid levels. Magnesium lowered the free cholesterol levels in the hearts of the rats on the cardiopathic diet.
5.1.3. Magnesium/Lipid/Catecholamine Interrelationships
Catecholamines have long been known to increase the blood levels of free fatty acids, whether by injection in animals (Dole, 1956; Bogdonoff et al., 1961) or as a result of such stress as myocardial infarction (Kurien and Oliver, 1966; Kurien et al., 1971; Oliver et al., 1968; Editorials, Lancet, 1969a,b). The complex interrelationships of magnesium and catecholamines and corticosteroids have been surveyed by Wallach (1976/1980) and those of corticosteroids (which are also released by stressful situations) and magnesium by Massry and Coburn (1973). In the case of the catecholamines, depending on the time of testing and the test situation, they have both increased and decreased blood magnesium levels and decreased tissue (i.e., heart) magnesium levels.
Considered here are the blood lipid-magnesium interrelationships, as influenced by cold-stress, the acute alcohol-withdrawal syndrome, and the administration of catecholamines, which might provide some insight into the somewhat contradictory findings. Rayssiguier and Larvor (1976/1980) have recently reported that either fasting or exposure of shorn young sheep to cold temperatures causes comparable lipolysis to that produced by infusions of either epinephrine or theophylline. Hypomagnesemia accompanied the increase in free fatty acid levels in the blood, caused by each of the stimuli. Sodium nicotinate, which is antilipolytic, inhibited both the increase in free fatty acids in the blood and the decrease in blood magnesium. High blood levels of long-chain free fatty acids are also seen during the acute phase of alcohol withdrawal (Mays et al., 1970) and the severity of the symptoms tends to be greater in those with higher levels of the fatty acids (Blink et al., 1973, 1976/1980). Since such fatty acids can chelate magnesium, Flink et al., (1976/1980) propose that the signs of alcohol withdrawal may depend upon inactivation of magnesium by the fatty acids. They verified the elevation of free fatty acids in the blood of dogs induced to imbibe alcohol and suggested that reducing lipolysis during alcohol withdrawal might be useful in controlling the symptoms, which are often controllable by magnesium repletion (Rink et al., 1954, 1957; Rink, 1956, 1969, 1976/ 1980)
The mobilization by catecholamines of free fatty acids and their inactivation of magnesium recall Browne's early (1964a,b) deduction that the clinical benefit reported from magnesium therapy of hyperlipemic patients with occlusive arterial disease might derive from magnesium-catecholamine interrelationships. He pointed out that magnesium inhibits catecholamine release from the adrenal medulla (Douglas and Rubin, 1961, 1963, 1964), and that smoking or nicotine infusions (to dogs) causes elevation of both serum free fatty acid levels and urinary excretion of catecholamines (Kershbaum and Bellet, 1964). The arrhythmia following clinical myocardial infarction might be related to the catecholamine-induced increase in circulating fatty acids that might be mediated by inactivation of serum magnesium. Perhaps more likely is the possibility that increased myocardial lipids, such as have been attributed to catecholamine lipid mobilization in rats injected with sympathomimetic agents (Ferrans et al., 1964, 1969) and in electrolyte-steroid cardiopathy (Prioreschi, 1966), might be the result of inactivation by the intramyocardial fats of cellular magnesium. It is provocative, in this regard, that a direct correlation was made by Balazs et al., (1962) with the cardiotoxicity of isoproterenal in rats and the amount of excess body fat. The availability of more fat for lipolysis under stressful situations might explain the greater susceptibility of obese individuals to fatal ischemic heart disease.
5.1.4. Estrogen, Lipids, and Magnesium; Interrelationships with Arteriosclerosis and Thrombosis
5.1.4.1. Estrogen Therapy of Ischemic Heart Disease
The treatment of men with coronary insufficiency by estrogens is no longer advocated. Estrogens used to be administered in an effort to lower the β/α-lipoprotein ratio of these men to that of young women (Ban, 1955; Oliver, 1960) because of the sex difference in the incidence of ischemic heart disease. Despite success in lowering the β- and raising the α-lipoprotein levels by giving estrogens (Barr et al., 1952; Townsend et al., 1952; Gertler et al., 1953, Steiner et al., 1955; R. W. Robinson et al., 1956; Voyles and Evans, 1961), there has not been satisfaction that a sufficiently suppressive effect is exerted on recurrence of cardiovascular accidents to justify the unpleasant side effects (Steiner et al., 1955; Oliver, 1962; Robinson et al., 1963).
The effect of estrogens on blood coagulation may provide a possible explanation of their failure to achieve benefit in patients who had suffered a myocardial infarction, a condition associated with increased coagulability of the blood shortly after the event (McDonald and Edgill, 1957, 1959; Katz et al., 1963). Estrogens have long been known to increase the coagulability of blood, an attribute that has been used to stop bleeding [e.g., after tonsillectomies (S. Fox, 1960) and to control epistaxis (E. Blackburn, 1963)]. This activity, however, seems relevant to the correlation of thromboses and infarctions with the use of estrogen-containing oral contraceptives (Inman et al., 1970; Coronary Drug Project Report, 1973a; Editorial, Lancet, 1977; Goldsmith and Johnston, 1979).
If one accepts the premise that formation of mural thrombi is a pathogenic mechanism in atherogenesis (Duguid, 1946; T. Crawford, 1959; Astrup, 1959; McDonald, 1959; Pilgeram, 1961; Pickering, 1963. A. Katz et al., 1963), the enhancement of intravascular coagulation by estrogens should result in a higher, rather than the lower incidence of cardiovascular disease in premenopausal women than in men.
5.1.4.2. Estrogen, Cardiovascular Effects, and Magnesium
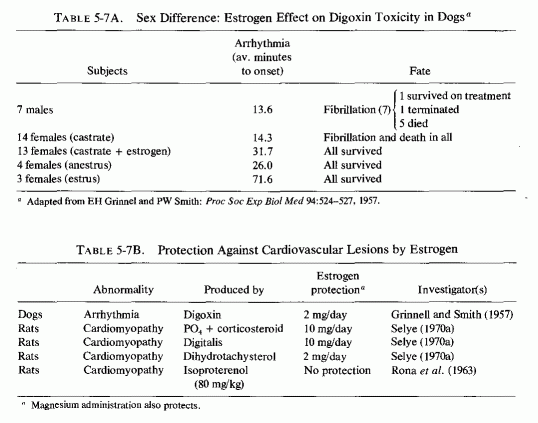
It was speculated (Seelig, 1964) that the common denominator between the low incidence of ischemic heart disease in men from the Orient (where the intake of saturated fat is low) and in young women might be magnesium. The substantially higher dietary intakes of magnesium in the Orient, and the better retention of magnesium by young women than young men on the customary marginally adequate magnesium intake of the Western world, suggested that the adequacy of magnesium might be the protective factor against IHD. If the effective retention of magnesium by women reflects its affinity, not only to target tissues such as those involved in the reproductive process and to bone (Walaas, 1950; Csapo, 1956; Best and Pickles, 1965; N. Goldsmith, 1971) but to the cardiovascular system, that might elucidate the greater resistance of premenopausal women than of older women and of both young and older men to cardiovascular disease. Thus far, experimental data verifying the greater resistance of females than males to cardiopathic agents have not elucidated the possible role of cardiovascular magnesium levels. One can draw some inferential conclusions, but definitive work remains to be done. For example, female dogs and rabbits are more resistant than are males to digitalis-induced arrhythmias (Grinnell and Smith, 1957; Rodensky and Wasserman, 1964). Castrated females are as susceptible as are males to digitalis toxicity; estrogen replacement (2 mg/day) markedly improves their resistance to arrhythmias, but not to the extent seen in estrus (Table 5-7A, Grinnel and Smith, 1957). High-dosage estrogen has been almost fully protective against the myocardial necrotic lesions produced by phosphate and corticoids and by digitalis overdosage and has protected against myocardial necrosis caused by dihydrotachysterol (Table 5-7B, Selye, 1970a). It is thus of interest that digitalis toxicity is increased by magnesium deficiency (Vitale et al., 1963), that digitalis increases the renal excretion of magnesium (Kupfer and Kosofsky, 1965), and that magnesium is useful in digitalis toxicity (Zwillinger, 1935; Szekely and Wynne, 1951; J. Stanbury and Farah, 1960; Cook et al., 1967; Wacker and Parisi, 1968; Seller et al., l970a,b). Although Rona et al. (1963) could not protect against the myocardial necrosis (produced by massive doses of isoproterenol) with estrogens, female rats are more resistant than are males to this form of cardiac damage, an effect that Rona et al. (1963) attributed to the slower rate of growth of the females. Possibly, the protective effect of estrogens in these experimental models might be mediated by increased uptake of magnesium by the myocardium, as well as by other tissues, in response to estrogen. Additional evidence that estrogen, or other female sex hormones, might be protective against several forms of cardiovascular lesions derives from study of the influence of pregnancy on experimental models. Advanced pregnancy in rats has protected against: (1) dihydrotachysterol-induced arteriosclerosis (Selye, 1957); (2) the cardiovascular necrosis and calcification and calcification of vitamin D excess (Potvliege, 1962); phosphate + corticoid-induced cardiomyopathy (Selye, 1958a); and hyperparathyroid myocardial necrosis (Lehr and Krukowski, 1961a,b; Krukowski, 1961, 1963; Lehr, 1965b). Pregnant dogs are more resistant than are nonpregnant females to necrotizing arteritis produced by a high-fat diet and renal insufficiency (Holman and Jones, 1953).
5.1.4.3. Magnesium, Estrogen, and Thrombotic Events
It is possible that the paradoxical effects of estrogen on diseases of the cardiovascular system relate partially to its effects on magnesium distribution. It has been shown that serum magnesium falls with the cyclic increase in estrogen secretion (Dahl, 1950; Nida and Broja, 1957; Goldsmith, 1963; Goldsmith et al., 1970; Goldsmith, 1971). The use of estrogen-containing oral contraceptives has been shown to reduce the serum levels of magnesium (in users versus nonusers) by 16% (Goldsmith et al., 1966), 28% (DeJorge et al., 1967), and by 27% and 33% (Goldsmith, 1971). Evaluation of different contraceptives suggests that it is the estrogen moiety that is responsible for the decrease in serum magnesium (Goldsmith and Goldsmith, 1966; Goldsmith et al., 1970, Goldsmith and Johnston, 1976/1980) although there are conflicting findings. Since rats given estrogen showed decreased serum magnesium levels, without increased urinary magnesium output, and since the bone-magnesium increased, Goldsmith and Baumberger (1967) proposed that a shift of magnesium to the tissues was responsible for the estrogen-induced fall in serum magnesium. Indirect support for the importance of the estrogen component of contraceptives in lowering serum magnesium comes from the report that progestogens increase rather than decrease serum magnesium (Dale and Simpson, 1972). Yet, norethisterone and mestranol, alone or combined, have been shown to increase magnesium levels in bone, muscle, and intestinal wall tissues (Gozan and Charnot, 1973; Charnot et al., 1974). Despite the increase in tissue levels of rats on mestranol, their serum magnesium levels did not fall; norethisterone, however, produced a 30% drop in serum magnesium (Gozan and Charnot, 1973). The picture is further confused by the studies showing no effect of several oral contraceptives on serum magnesium (N. Hahn et al., 1972) or on magnesium levels of plasma, erythrocytes, and platelets (Thin, 1971). Data on decreased serum magnesium levels during pregnancy are discussed elsewhere in this volume, as possibly reflecting a true magnesium deficit rather than a hemodilution or estrogen-induced effect. Wallach (1976/1980) has considered the findings relating to the effect of estrogen on magnesium and has commented that circumstantial evidence from studies of interrelations of estrogen, calcium, and magnesium on thymic cell proliferation (Morgan and Perris, 1974) suggests that estrogen may favor cellular transport of magnesium.
Although there is no uniform agreement that estrogens lower serum magnesium levels, most of the evidence points in that direction. Thus, the still controversial evidence that low magnesium levels can contribute to coagulopathy deserves consideration as a possible factor in estrogen-induced thrombotic disorders. Durlach (1967a,b,c) first described severe thromboembolic disease in a young woman with latent tetany of magnesium deficiency. Her disorder was associated with increased ADP-induced platelet aggregation. Additional instances have since been reported in women with latent tetany of magnesium deficiency (DuPont et al., 1969; Durlach, 1970; Boudet et al., 1972; Erödi, 1973; Debrand, 1974; Maurat et al., 1974; Seelig et al., 1976/1980). Durlach (1970) has also shown that estrogen therapy gives rise to both functional platelet alterations and to signs of magnesium deficiency, which regress on administration of oral magnesium in moderate dosage. Vajna (1971/1973) has claimed that administration of magnesium to women on oral contraceptives significantly reduces the risk of coagulopathy.
Elin (1976/1980) and Durlach (1976/1980) have reviewed the in vitro evidence that magnesium plays a role (predominantly inhibitory) in the coagulation process. However, as Durlach (1976/1980) stresses, most of the in vitro studies showing that magnesium can inhibit coagulation factors-prothrombin, thrombin, and Factors V, VII, and IX-and can increase fibrinolysis, have been based on studies with high magnesium concentrations. They are thus not directly relevant to consideration of the effects of low or marginally low serum magnesium levels on the tendency toward intravascular coagulation. A few experimental magnesium-deficiency studies may shed light on the clinical coagulopathy of magnesium deficiency or on that accompanying use of agents (such as estrogens) that lower serum magnesium levels. Stevenson's and Yoder's (1970) magnesium-deficient animals had significantly shorter thrombin clotting time and greater ADP breakdown than did the normal group (p = 0.001). The partial thromboplastin time was also significantly reduced in magnesium-deficient calves (p = 0.05). Stachura (1971) observed hypercoagulation with shortened thromboplastin time in magnesium-deficient rats. Magnesium-deficient calves had insignificantly increased ADP-platelet aggregation (p = 0.05) but magnesium-deficient rats had more ADP-platelet aggregation (p = 0.05) than did normal rats (Stevenson and Yoder, 1970). Since magnesium-deficient rats commonly develop hypercalcemia (Larvor and Durlach, 1971a; Seelig and Haddy, 1976/1980),the species difference in ADP platelet response to magnesium deficiency might reflect the presumed difference in the Mg/Ca ratio in the rats versus the calves (data on calcium levels were not provided).
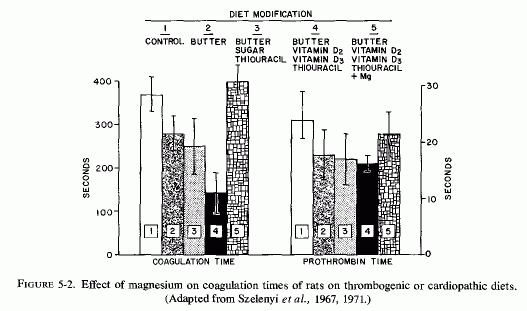
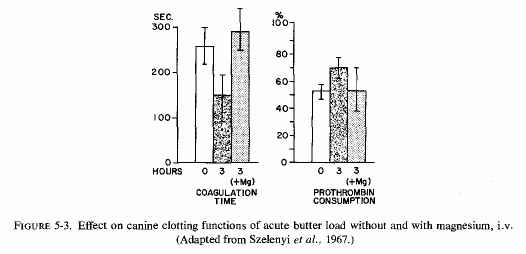
Hypercoagulability, produced by feeding rats a thrombogenic diet containing cholesterol, vitamins D2 and D3 and thiouracil, as well as large amounts of butter, was counteracted by oral magnesium chloride (Szelenyi et al., 1967) (Fig. 5.2). Dogs acutely loaded with butter showed markedly increased blood coagulability three hours later. Magnesium, given intravenously at the time of intragastric butter administration, prevented the decreased coagulation time and the increased prothrombin consumption (Szelenyi et al., 1967) (Fig. 5-3).
Clinical studies of the effects of magnesium administration to patients with cardiovascular disease and hyperlipidemia have been considered. Relevant to this section are the reports that parenteral magnesium therapy reduced plasmin inhibition (R. Parsons, 1958). Other studies suggest that magnesium therapy accelerates fibrinolysis (Hackethal, 1949; Zahnert and Oloffs, 1960). Further clinical investigation of effects of magnesium on blood coagulation and clot lysis is required.
Elin (1976/ 1980) has reviewed in vitro evidence that magnesium affects platelet aggregation and release. Its inhibitory effects on platelet aggregation have been with high concentrations (Born and Cross, 1964); the calcium/magnesium ratio is important at low concentrations (Herrman et al., 1970). Platelet release is calcium dependent (Sneddon and Williams, 1973); increasing concentrations of magnesium are inhibitory (Sneddon, 1972).
Whether these findings are relevant to the increased blood coagulability and platelet adhesiveness of patients with myocardial infarctions remains to be resolved. Hughes and Tonks (1965) reported significantly decreased serum magnesium levels and increased platelet aggregability in infarct patients, as compared with matched controls, a finding reported also by Prakash et al. (1971/1973). Chadda et al. (1976/1980) have also reported decreased serum magnesium in such patients. However, Murnaghan et al. (1969) reported elevated serum magnesium levels and Khan et al. (1974) normal levels, the latter in association with highly significantly increased platelet adhesiveness. In view of the known stress and anoxia-induced magnesium egress from the tissues, with initially increased serum magnesium, followed later by decreased serum magnesium levels, longitudinal studies of infarction patients must be done with meticulous attention paid to the time lapse after the ischemic event; and to the degree of decompensation-hypoxia.
(include the word "jacket" to search only in this book)
| Jacket | Preface | Contents | Introduction (Chapter 1) |
Chapter: | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Appendix | Bibliography (A-D), (E-K),
(L-R), (S-Z) |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}